Typus der Dorfgemarkung
Kulturlandschaft lässt sich nur verstehen in Beziehung zur historischen Landwirtschaft und damit zur bäuerlichen Siedlung, dem Dorf. Somit ist die Dorfgemarkung die Grundeinheit der Kulturlandschaft. Für die nordwestdeutsche Dorfgemarkung, die hier beispielhaft dargestellt werden soll, sind zwei Gradienten wesentlich, die sich überlagern und die Vielfalt an Pflanzengesellschaften bedingen. Der bäuerliche Kulturgradient bringt Zonen unterschiedlicher Kulturintensität hervor, wobei die intensiv bewirtschafteten Flächen in Dorfnähe und die extensiv genutzten Bereiche am äußeren Rand der Gemarkung liegen. Vorwiegend durch die Düngung und den Stoffentzug durch Ernte entsteht auf diese Weise ein Gefälle von nährstoffreichen Flächen in Dorfnähe bis zu nährstoffarmen Flächen in der Außenzone der Gemarkung.

Der zweite Gradient ist von Ellenberg herausgearbeitet worden. Er fand heraus, dass das historische, also landwirtschaftlich orientierte Dorf in Nordwestdeutschland immer auf der Grenze zwischen einem feuchten und einem trockenen Landschaftsraum angelegt wurde. Damit ist eine Aufteilung der Dorfgemarkung in einen feuchten und einen trockenen Bereich vorgegeben. Durch die Überlagerung dieses Feuchtegradienten mit dem Kulturgradienten ergibt sich die prinzipielle Vegetationsgliederung der nordwestdeutschen Dorfgemarkung. Sie soll im folgenden hinsichtlich der Vegetationsgestalten dargestellt werden.
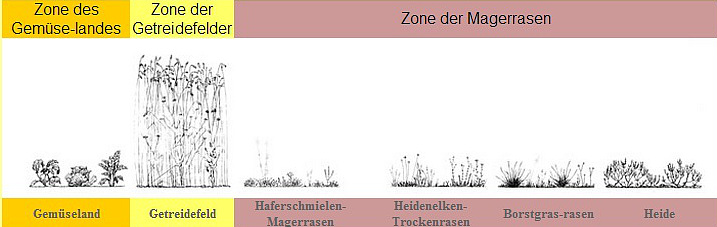
Im trockenen Wirtschaftsraum gliedert sich der Kulturgradient in die drei Zonen der Magerrasen, der Getreidefelder und des Gemüselandes. Die Magerrasen sind durch niedrige Wuchshöhe und das Vorherrschen strahliger Pflanzenformen gekennzeichnet. Die ohnehin linealische Form der Gräser wird in dieser Gesellschaft durch die Ausbildung nadeliger Formen ins Extrem getrieben (z.B. bei Borstgras und Schafschwingel). Auch bei vielen Kräutern zeigt sich die linealische Gestalt in Form von nadeligen oder linealischen Blättern, flächige Blätter sind zumeist in Bodennähe als Rosette ausgebildet. Die Magerrasen zeichnen sich durch starke Blühaspekte aus.
In der Zone der Kornfelder sind die linealischen Pflanzengestalten zwar vorherrschend, machen jedoch im Jahreslauf eine deutliche Metamorphose durch. Im Herbst oder Frühjahr, nach dem Austrieb des Getreides, sind dessen Blätter verhältnismäßig breit und kräftig grün, so dass die Vegetationsgestaltung zu dieser Zeit mehr in flächige Formen geht. Ganz anders stellt sich die Situation im Sommer zur Reifezeit des Getreides dar. Dann ist jede Ähnlichkeit mit flächenblättrigen Gestalten verschwunden, und zu der rein linealischen Form gesellen sich die Körner in einer darüber angeordneten Schicht.
Im Gemüseland schließlich herrschen flächige Gestalten vor. Strahlige Formen oder Blühaspekte werden durch die Bearbeitung unterdrückt, da die gesamte Biomasse in den Sprossbereich (bzw. Wurzelbereich) investiert werden soll. Eine Jahreslauf-Metamorphose, vergleichbar mit jener der Getreidefelder, existiert im Gemüseland nicht; hier gibt es nur die ständige Zunahme an Masse ohne gravierende Formänderung.
Im Gestaltvergleich aller drei Zonen zeigt sich eine durchgehende Metamorphose-Richtung. Die Magerrasen sind durch strahlige Formen und starke Blütenbildung gekennzeichnet. Zum Getreidefeld hin nimmt die Strahligkeit insofern ab, als sie hier auf den Sommer beschränkt ist, und Blühaspekte treten vor allem in Randzonen der Felder auf. Im Winterhalbjahr zeigt sich eine mehr flächige Gestaltung. Diese wird dann im Gemüseland vorherrschend, und die Blütenbildung verschwindet ganz. Die Gestaltreihe zeigt also eine starke Polarisierung in Magerrasen und Gemüseland, wobei das Getreidefeld zwischen diesen beiden Extremen vermittelt, und zwar im zeitlichen Rhythmus.
Ein ebenfalls zeitlicher Rhythmus betrifft die Nutzung der Kornfeld-Zone, indem hier in bestimmter Abfolge Getreide und Hackfrüchte kultiviert werden und manchmal ein beweidetes Brachestadium eingeschoben wird (z.B. Feld-Gras-Wirtschaft). Dieser Wechsel bedeutet, dass die Zone der Kornfelder bei Hackfruchtanbau dem Gemüseland ähnlich wird und bei Brache und Beweidung Magerrasencharakter bekommt. Nur bei Getreideanbau zeigt sie ihr eigentliches Gesicht. Gerade dieses Pendeln zwischen den Charakteren der innersten und äußersten Zone der trockenen Dorfgemarkung zeigt sowohl die räumliche als auch funktionelle Vermittlung zwischen beiden.
Im feuchten Teil der Dorfgemarkung können folgende Zonen unterschieden werden: Zone der mageren Feuchtwiesen, Zone der Fettwiesen, Zone der Fettweiden. Die mageren Feuchtwiesen setzen sich je nach Wasserhaushalt aus verschiedenen Pflanzengesellschaften zusammen, z.B. Binsen-Pfeifengraswiesen und Kleinseggenrieden, sowie einigen weiteren, kleinflächig eingestreuten Gesellschaften, beispielsweise die Waldbinsen-Wiese auf Sickerquell-Standorten.

Hinsichtlich der Vegetationsgestalt sind sich Binsen-Pfeifengraswiese und Kleinseggenried recht ähnlich. Beide sind niedrigwüchsig und vom linealischen Gestaltelement beherrscht, wobei flächenblättrige Formen trotz nassem Wuchsort überraschend wenig vertreten sind. Insofern sind sie den Magerrasen des trockenen Wirtschaftsraumes nicht unähnlich, und entsprechend gibt es zwischen den trockenen Magerrasen und den mageren Feuchtwiesen viele Übergänge. Von den frischgrünen Fettwiesen und -weiden setzen sich die mageren Feuchtwiesen deutlich farblich ab: Die Binsen-Pfeifengraswiese weist ein graues Blaugrün auf, während beim Kleinseggenried mehr bräunliche Töne vorherrschend sind.
Die Zone der Fettwiesen wird, je nach Wasserhaushalt, geprägt durch Glatthafer-Wiesen und Sumpfdotterblumen-Wiesen. Beide sind recht hochwüchsig, besonders im Vergleich zu den mageren Feuchtwiesen. Sie zeigen deutliche Schichtungen in eine Oberschicht aus linealischen Strukturen und eine von flächenblättrigen Elementen gebildete Unterschicht. Letztere ist besonders bei der Sumpfdotterblumen-Wiese sehr ausgeprägt.
In der Zone der Fettweiden ist die Weidelgras-Weißklee-Weide die kennzeichnende Gesellschaft, die stellenweise verzahnt ist mit dem Breitwegerich-Trittrasen an stärker betretenen Stellen. Die Weidelgras-Weißklee-Weide als zonenprägende Gesellschaft ist sehr niedrigwüchsig und ähnelt damit den mageren Feuchtwiesen. Anders als diese ist die Weide jedoch in ihrer Unterschicht sehr stark von flächigen Gestalten beherrscht, die als mehr oder weniger dichte Blätterdecken von Weißklee, Kriechhahnenfuß u.a. auftritt. Ebenso deutlich ist der Farbunterschied: zu den rötlichen und bläulichen Tönen der feuchten Magerwiesen steht das ganzjährige frische Gelbgrün der Weide im scharfen Gegensatz. Auffällige Blühaspekte findet man selten, da vorher alles abgefressen wird.
Es zeigt sich also, dass wir es sowohl im trockenen wie im feuchten Bewirtschaftungsraum im Grunde mit jeweils drei deutlich voneinander unterschiedenen Vegetations-Gestalttypen zu tun haben. Sowohl für die Reihe der trockenen als auch der feuchten Seite der Dorfgemarkung erkennt man eine Gesetzmäßigkeit in Form einer Polarität zwischen einem dorfnahen Bereich mit üppigem Wachstum, reicher Blattentfaltung und Blütenarmut und einem dorffernen Bereich mit reduziertem Wachstum, gehemmter Blattentfaltung, strahligen Formen und auffälligen Blühaspekten andererseits. Die Mitte zwischen diesen beiden Polen wird gehalten durch eine Gestalt, die sowohl durch ihre Wuchshöhe auffällt als auch Elemente des einen wie des anderen Pols in sich vereinigt. Im Getreidefeld ist dies vor allem durch den jahreszeitlichen Rhythmus gegeben, indem es im Frühjahr mehr zum vegetativen Pol des Gemüselandes tendiert und im Sommer zur Reifezeit den Trockenrasen ähnlich wird. In der Wiese überlagern sich die beiden Pole mehr räumlich: Die breitblättrige Unterschicht stellt die Beziehung zu der Weidelgras-Weißklee-Weide her, die feinblättrige Oberschicht mit den Blüten ist mehr mit den Magerwiesen (sowohl trockener als auch feuchter Standorte) verwandt.
Eigene Publikationen zum Thema
- H.-Ch. Vahle (1991): Die Idee der Kulturlandschaft – Am Beispiel Nordwestdeutschlands. – Die Drei 61 (7/8): 581-612.
- H.-Ch. Vahle (1997): Zum Typus der nordwestdeutschen Kulturlandschaft. – Natur- und Kulturlandschaft 2: 102-112.
- H.-Ch. Vahle (1997): „Organismus Landschaft“ – mehr als ein Ökosystem? – Jahrbuch der Baumpflege 1997: 78-97.
- H.-Ch. Vahle (1998): Auf der Suche nach der Leitidee der nordwestdeutschen Kulturlandschaft. – In: Suchantke, A. (Hrsg.): Goetheanistische Naturwissenschaft Band 5: Ökologie. Stuttgart: 312-368.
- H.-Ch. Vahle (2000): Die Vegetationsästhetik der Kulturlandschaft. – In: Pedroli, E. (ed.): Landscape – Our Home. Lebensraum Landschaft. – Essays über die Kultur der europäischen Landschaft als Aufgabe: 179-186. Zeist, Stuttgart.
- H.-Ch. Vahle (2001): Das Konzept der potentiellen Kulturlandschafts-Vegetation. – Tuexenia 21: 273-292. Göttingen.
- H.-Ch. Vahle & Y. Zimmermann (2002): Steigerung der Biodiversität in einer vorwiegend agrarisch genutzten Landschaft südlich des Ruhrgebietes. – In: „Treffpunkt biologische Vielfalt II“ (Hrsg.: Bundesamt für Naturschutz): 151-156. Bonn.
Kulturlandschaft als Organismus
Durch den Gestaltvergleich der Vegetation innerhalb der Dorfgemarkung wird eine Ordnung ersichtlich, die eine innere Stimmigkeit vermuten lässt und die als ein erster Hinweis auf eine organische oder sogar organismische Ganzheit gewertet werden kann. Wie kann aber die organismische Ganzheit oder der Organismus begrifflich so gefasst werden, dass sie über die individuellen Erscheinungen von Mensch, Tier und Pflanze hinaus anwendbar werden?
Die historische Dorfgemarkung ist in der Literatur vielfach als ein fein abgestimmter Funktionszusammenhang beschrieben worden, jedenfalls aus der Sicht einer ökologisch nachhaltigen Nutzung. Nun ist vielleicht überraschend, dass sich auch in der Vegetation ein ebenso fein abgestimmter Gestaltzusammenhang zeigt, der mit dem Funktionszusammenhang parallel geht. Die Prozesse der Landbewirtschaftung, die dadurch veränderten und neu organisierten ökologischen Systeme, die unter der Nutzung neu entstehenden Biotope und schließlich die Vegetationsgestalt bilden solch ein harmonisches Ganzes, dass sich die Frage aufdrängt, ob man hierbei womöglich sogar von einem Organismus höherer Ordnung sprechen kann.
Ausgehend vom menschlichen Organismus entwickelte Rudolf Steiner (1917) einen Organismus-Begriff, der so allgemein gehalten und flexibel ist, dass er prinzipiell auch auf andere lebendige Komplexe angewandt werden kann. Er fand eine Gliederung des menschlichen Organismus in drei Organsysteme, die er Stoffwechsel-Gliedmaßen-System, rhythmisches System und Nerven-Sinnes-System nannte. Ihr Funktionszusammenhang ist folgender: Durch das Stoffwechsel-Gliedmaßen-System, hier abgekürzt Stoffwechsel-System genannt, baut der Organismus körpereigene Substanz auf und schafft damit seine Lebensgrundlage. Es ist gekennzeichnet durch intensive Wachstumskräfte; das Gewebe dieser Organe wird in bestimmten Abständen vollständig erneuert, die Reproduktionskraft ist hoch (Beispiele: Verdauungsorgane, Fortpflanzungsorgane).
Durch das Nerven-Sinnes-System bildet der Organismus die Umgebung in sich ab; er nimmt äußere Sinnesreize auf, die nicht „nur“ zu einem bestimmten Verhalten führen, sondern auch teilweise direkt in lebenswichtige physiologische Wirksamkeit umgewandelt werden, z.B. die körpereigene Herstellung von Vitamin D durch Lichteinwirkung. Das Gewebe dieses Organsystems wird, einmal gebildet, nicht mehr oder kaum mehr erneuert, die Reproduktionsfähigkeit und das Wachstum sind im erwachsenen Organismus äußerst gering (Beispiele: Sinnesorgane, Gehirn). Die Reduktion der Vitalkräfte ist notwendig für die Sinnes- und Nervenorganisation. Das Auge beispielsweise kann seine Funktion nur erfüllen, weil in wichtigen Teilen die Stoffwechselprozesse ganz zurückgenommen sind. Man muss sich einmal vorstellen, was das Auge wäre, wenn Linse und Glaskörper von feinen Blutgefäßen durchzogen würden: dann würden wir nichts mehr sehen können! Das Auge muss, um als Sinnesorgan dienen zu können, viel weniger lebendig-vital sein als die Stoffwechselorgane der Bauchhöhle.
Zwischen beiden Systemen vermittelt das rhythmische System durch die Tätigkeit des Kreislaufes der Körperflüssigkeiten (vor allem Blutkreislauf) und der Atmung. Stoff- und Gasaustausch zwischen Organen und zwischen Organismus und Umwelt wird hierdurch gewährleistet. Die Tätigkeit dieser Organe ist eine stark rhythmische (periodische): Herzschlag, Atmung.
Durch die Idee des dreigliedrigen Organismus lieferte Steiner die allgemeine Begrifflichkeit, mit der auch die Gliederung der Dorfgemarkung organismisch verstanden werden kann. Danach werden die drei Dorfgemarkungs-Zonen auf einer ganz allgemein-funktionellen Ebene mit den drei Organsystemen des Menschen vergleichbar. Es lassen sich so Parallelen finden zwischen der äußeren Gemarkungszone (feuchte und trockene Magerrasen) und den Sinnesorganen sowie zwischen der inneren Zone (Gemüseland und Fettweiden) und dem Stoffwechsel-System. Beim Vergleich von mittlerer Zone (Kornfelder und Wiesen) und dem rhythmischen System stößt eine einfache Gegenüberstellung auf Schwierigkeiten. Dennoch ist das gemeinsame Charakteristikum beider Systeme die Vermittlung zwischen den beiden Polen, und zwar auf eine rhythmische Art und Weise.

Eigene Publikationen zum Thema
- H.-Ch. Vahle (1991): Die Idee der Kulturlandschaft – Am Beispiel Nordwestdeutschlands. – Die Drei 61 (7/8): 581-612.
- H.-Ch. Vahle (1997): Zum Typus der nordwestdeutschen Kulturlandschaft. – Natur- und Kulturlandschaft 2: 102-112.
- H.-Ch. Vahle (1997): „Organismus Landschaft“ – mehr als ein Ökosystem? – Jahrbuch der Baumpflege 1997: 78-97.
- H.-Ch. Vahle (1998): Auf der Suche nach der Leitidee der nordwestdeutschen Kulturlandschaft. – In: Suchantke, A. (Hrsg.): Goetheanistische Naturwissenschaft Band 5: Ökologie. Stuttgart: 312-368.
- H.-Ch. Vahle (2020): Die Kulturlandschaft als Organismus und ihr Entwicklungspotenzial. – In: Rosslenbroich, B. (Hrsg.): Perspektiven einer Biologie der Freiheit. Autonomieentwicklung in Natur, Kultur und Landschaft. Verl. Freies Geistesleben, Stuttgart: 450-489.
- H.-Ch. Vahle (2020): Lichtlandschaft – die Zukunft der Kulturlandschaft? – Die Christengemeinschaft 92 (7-8): 18-21. Stuttgart
Potenzielle Kulturlandschafts-Vegetation
Das Vegetationspotenzial der Landschaft
Die Pflanzengesellschaften Mitteleuropas sind zum größten Teil durch die historische Landschaftskultur entstanden. Die Vielfalt an Biotopen und Arten hat damit zugenommen. Ohne den Menschen würde die Erde hier überwiegend von Wald bedeckt sein. Die menschbedingte Vielfalt ist jedoch nichts Willkürliches; auch die neu entstandenen Pflanzengesellschaften sind charakteristisch für die jeweilige Landschaft.
Dass heutzutage längst nicht mehr alle Pflanzengesellschaften in „ihren“ Landschaften verwirklicht sind, liegt unter anderem daran, dass die entsprechende Vielfalt der nachhaltig-ökologischen Nutzungsmöglichkeiten nicht mehr realisiert wird. Demnach ist das Potenzial, also die prinzipielle Möglichkeit, noch vorhanden und könnte bei der Wiederaufnahme oder Neuentwicklung geeigneter Nutzungen wieder Wirklichkeit werden. Die Gesamtheit der in einem Gebiet möglichen Pflanzengesellschaften, die durch die Nutzung des Menschen prinzipiell möglich sind, nennen wir die Potenzielle Kulturlandschafts-Vegetation (PKV).
Der Natürliche Wald
… und was durch den Menschen daraus werden kann.




Die meisten Pflanzengesellschaften hatten einen ganz bestimmten Platz im Nutzungsmuster der historischen Landbewirtschaftung, das durch die Struktur der Dorfgemarkung vorgegeben wurde (siehe 1. Kapitel „Typus der Dorfgemarkung“). Die Überlagerung von Kulturgradient und Feuchtegradient bedingt hierbei die Vielfalt an anthropogenen Vegetationstypen.
Das Vegetationspotential der Gemarkungszonen: Beispiel Weidezone
Jede der 6 Gemarkungszonen beinhaltet neben der großflächig auftretenden, namengebenden Pflanzengesellschaft noch eine große Zahl weiterer Vegetationstypen. Beispielhaft sei die Zone der Fettweiden etwas genauer dargestellt.

Da ist zunächst die Weidelgras-Weißklee-Weide (Lolio-Cynosuretum) als die typische Weidegesellschaft, die in ihrer Artenkombination dadurch erhalten wird, dass sie von Weidetieren abgefressen wird.
Unvermeidbar sind in solchen Weiden Trampelpfade des Viehs und andere Stellen, wo die geschlossene Grasnarbe durch Tritt zerstört ist. Hier wird der Weiderasen durch den Breitwegerich-Trittrasen (Lolio-Plantaginetum) ersetzt. Ihn findet man auch an den Weidetoren, wo er den offenen, zertretenen Boden kranzförmig umgibt.



Dieser offene Boden wiederum ist Lebensraum der Vogelknöterich-Wegkamillen-Gesellschaft (Polygono-Matricarietum discoideae), eines einjährigen Trittrasens, der noch stärkere Störungen als der Breitwegerich-Trittrasen verträgt. Bei genügend Feuchtigkeit findet man im Frühling an den gleichen Stellen manchmal eine ganz spezielle einjährige Trittrasen-Gesellschaft, den Mäuseschwänzchen-Rasen (Myosuretum minimi). Während die häufige Vogelknöterich-Wegkamillen-Gesellschaft nicht nur in Weiden, sondern praktisch an allen betretenen Stellen wächst, ist der Mäuseschwänzchen-Rasen eine sehr typische Gesellschaft der feuchten Weideeingänge.

Da die Fettweide im feuchten Wirtschaftsraum liegt, kann auch das Grundwasser stellenweise dicht unter der Oberfläche stehen. Sind in der Weide etwas tieferliegende Senken vorhanden, steht hier das Wasser im Winterhalbjahr sogar über dem Boden, indem es vorübergehende Pfützen und flache Tümpel bildet.
Im Sommer ist das Wasser längst wieder verschwunden, und auf den ersten Blick bemerkt man gar nicht, dass hier andere Arten zusammen wachsen als in der angrenzenden Weidelgras-Weißklee-Weide. Da die Vegetation der Mulden im Winter überflutet wird, nennt man sie Flutrasen, und die kennzeichnende Pflanzengesellschaft ist der Knickfuchsschwanz-Rasen (Ranunculo repentis-Alopecuretum geniculati).
Überhaupt sind viele feuchteliebende Gesellschaften in der Weidezone vertreten. An besonders nassen Stellen, aber auch an anderen Orten, die sich als praktisch erwiesen haben, wurden Weidetümpel als Tränke für das Vieh angelegt. Durch die stark gedüngte Umgebung sind auch sie nährstoffreich, und entsprechend finden sich hier nährstoffliebende Pflanzengesellschaften.

Besonders charakteristisch sind die verschiedenen Wasserlinsen-Gesellschaften (Lemnetea), die die Wasserfläche manchmal ganz als „Entengrütze“ überziehen. Die sommerlich trocken fallenden Schlammbänke bedecken sich im Hochsommer mit kniehohen Krautfluren einjähriger Arten, die zur Wasserpfeffer-Zweizahn-Gesellschaft (Polygono-Bidentetum) gehören. Die Ufer werden von mittelhohen Röhrichten frischgrüner Arten gesäumt, vor allem vom Wasserschwaden-Röhricht (Glycerietum maximae) und Kalmus-Röhricht (Acoretum calami).


Schließlich wachsen an allen Säumen, also an Weiderändern, Grabenböschungen, Wegrändern, zwischen Weiderasen und Gebüschen und an anderen Grenzlinien verschiedene Staudengesellschaften. Auch sie lieben nährstoffreiche, feuchte Böden. Dazu gehört die häufige Brennessel-Giersch-Gesellschaft (Urtico-Aegopodietum), die auch zum Leidwesen vieler Gärtner nicht nur in der Weidezone wächst, weiterhin vor allem an Grabenböschungen die im Hochsommer rosa blühende Wasserdost-Gesellschaft (Eupatorietum cannabini) und die Rauhhaarweidenröschen-Zaunwinden-Gesellschaft (Epilobio hirsuti-Convolvuletum).
Auch Gebüsch-Gesellschaften fehlen nicht, dabei handelt es sich zumeist um artenärmere Ausbildungen des Weißdorn-Schlehen-Gebüsches (Crataego-Prunetum), in denen die Schlehe etwas zurücktritt und dafür Holunder und Salweiden häufiger sind.
Diese Pflanzengesellschaften sind nur einige Beispiele aus einer einzigen Zone, und zwar einer nicht gerade sehr vielfältigen Zone der Kulturlandschaft. Man kann sich vielleicht denken, wie groß die Zahl der möglichen Gesellschaften in der gesamten Kulturlandschaft des eng begrenzten Gebietes einer Dorfgemarkung ist! Und wie wenig wird darüber gewusst, welche Gesellschaft sich bei welcher Nutzung einstellt bzw. welche Nutzung zu welcher Gesellschaft gehört! In der Vegetationskunde sind diese Zusammenhänge zwar bekannt, aber kaum für die Praxis aufbereitet und allgemeinverständlich beschrieben. Da besteht großer Nachholbedarf.
Vegetations-Leitbilder
Mit der potenziellen Kulturlandschafts-Vegetation ist ein großer Fundus an Gestaltungsmöglichkeiten gegeben, der für die Entwicklung landschaftlicher Leitbilder eine Bedeutung hat. Dabei ist die Neugestaltung des Dorfes und der Dorfgemarkung der Punkt, an dem die Leitbilddiskussion um die Kulturlandschaft ansetzen muss. Es geht dabei um den Versuch, Landwirtschaft, Erholung und Naturschutz auf engem Raum so zu verbinden, dass sie sich nicht nur gegenseitig dulden, sondern einander fördern.
Mit dem Konzept der PKV liegt eine wichtige Grundlage für ein solches Leitbild vor. Sowohl im Sinne einer nachhaltigen Landbewirtschaftung als auch unter dem Aspekt der Erhaltung und Entwicklung von Arten- und Biotopdiversität ist es notwendig, in der Kulturlandschaft einen Trophiegradienten nach dem Vorbild des traditionellen Kulturgradienten wieder neu aufzubauen. Dadurch erhalten sowohl die verschiedenen Magerbiotope als auch die nährstoffreicheren, produktiven Agrarflächen (auch ökologisch bewirtschaftete Flächen sind eutroph) sowie die dörfliche Ruderalvegetation ihren angemessenen Platz in der Landschaft.
Der Kulturgradient der Dorfgemarkung mit seinen vegetationskundlich reich gegliederten Zonen kann dabei Vorbild für die Anordnung von Pflanzengesellschaften zu besserer (nachhaltiger, effektiver) Pflege und Entwicklung sein. Zonentypische Pflanzengesellschaften sollten demnach in unmittelbarem räumlichen Kontakt zueinander liegen oder gelegt werden.
Publikationen zum Thema
- H.-Ch. Vahle (2000): Die Vegetationsästhetik der Kulturlandschaft. – In: Pedroli, E. (ed.): Landscape – Our Home. Lebensraum Landschaft. – Essays über die Kultur der europäischen Landschaft als Aufgabe: 179-186. Zeist, Stuttgart.
- H.-Ch. Vahle (2001): Das Konzept der potentiellen Kulturlandschafts-Vegetation. – Tuexenia 21: 273-292. Göttingen.
- Y. Zimmermann (2004): Die potentielle Kulturlandschafts-Vegetation im Bergisch-Märkischen Hügelland. – Wiss. Schr.-R. Institut Evolutionsbiol. 10. Martina-Galunder-Verl. Nümbrecht.
Landwirtschaft und Vegetationsvielfalt
Die biologische Vielfalt einer Landschaft zeigt sich in der Vielfalt verschiedener Pflanzengesellschaften; die Pflanzengesellschaften sind sozusagen das Maß und das Gestaltungsmittel der biologischen Vielfalt.
Um eine Vorstellung von der lebendigen Vielfalt unserer Heimatlandschaften zu gewinnen, kann auf die Vegetationskunde zurückgegriffen werden. Für Deutschland sind gegenwärtig etwa 800 Pflanzengesellschaften beschrieben, was unter anderem die unglaubliche Differenzierung der heimischen Vegetation vor Augen führt. Von dieser Summe sind etwa zwei Drittel (also ca. 550) solche Pflanzengesellschaften, die irgendwie vom Menschen abhängig sind, die durch die Tätigkeit des Menschen entstanden sind. Das heißt nichts anderes, als dass die Vielfalt großenteils in den Händen von uns Menschen liegt, was gar nicht oft und deutlich genug gesagt werden kann.
Diese 550 Pflanzengesellschaften sind auch weiterhin auf Bewirtschaftung und Pflege oder sonstige, z.B. zufällige Einflüsse von uns Menschen (Betreten, Befahren …) angewiesen. Viele sind mehr oder weniger häufige „Allerwelts“-Pflanzengesellschaften, die auch unter den heutigen Bedingungen immer wieder spontan entstehen oder erhalten werden (z.B. Breitwegerich-Trittrasen an Wegrändern, Brennnessel-Giersch-Gesellschaften an Zäunen und Grabenrändern). Andere sind selten, da auf historische Nutzungsformen angewiesen: Feuchtwiesen, Magerwiesen, Trockenrasen, Heiden und viele andere. Für alle diese Pflanzengesellschaften und damit für die davon abhängigen Lebensgemeinschaften trägt die Landwirtschaft die Verantwortung.



In Anbetracht dessen, dass es vor allem die historische Landbewirtschaftung war, die diese Vielfalt hervorgebracht hat, kann man sich fragen, was denn die heutige Landwirtschaft zum Erhalt und zur Entwicklung von Vegetationsvielfalt beitragen kann. Die Frage ist auch, welche Form von Landwirtschaft das sein müsste. Der Ökolandbau bietet sicherlich die beste Grundlage, da hier nicht nur pestizidfrei und mit mäßiger, überwiegend organischer Düngung gearbeitet wird, sondern weil die Förderung von lebendiger Vielfalt zur Philosophie des Ökolandbaues prinzipiell dazugehört.
Eine Garantie für Vegetationsvielfalt ist der Ökolandbau allein jedoch noch nicht. Die bunte Ackerwildkraut-Vegetation wird durch ihn zwar noch am ehesten gefördert, aber im Grünland gibt es oft kaum Unterschiede zur konventionellen Landwirtschaft, von vielen anderen „Grenzertragsstandorten“ ganz zu schweigen. Das ist in vielen Fällen eine Frage der Finanzierung, aber auch des Bewusstseins.
Hier kann die Pflanzensoziologie Hilfestellung geben. Sie hat längst ein detailliertes Zusammenhangwissen erarbeitet, das für Projekte und Maßnahmen zur Steigerung von Vielfalt verwendet werden kann. Um durch gestaltende, pflegende und wirtschaftende Maßnahmen am jeweiligen Ort eine große Vegetations-Vielfalt mit der regionaltypischen Eigenart (lokale Vegetationstypen) zu entwickeln, ist die genaue Kenntnis der dort vorhandenen und möglichen anthropogenen Pflanzengesellschaften notwendig. Das Spektrum dieser Gesellschaften zu erarbeiten und für die Berater und die Bewirtschafter aufzubereiten, um die Biodiversität in der Agrarlandschaft zu steigern, sehe ich als eine besonders dringende Aufgabe der modernen angewandten Pflanzensoziologie an.
Zur Darstellung der Vegetations-Vielfalt einer Region beschränke ich mich nicht auf die aktuell vorhandenen Pflanzengesellschaften, sondern beziehe alles das mit ein, was früher hier einmal vorkam und was unter bestimmten (zur Zeit nicht verwirklichten) Nutzungsbedingungen vorkommen könnte. Alles das – vergangene, gegenwärtige und zukünftig mögliche Pflanzengesellschaften – ist die Potenzielle Kulturlandschafts-Vegetation einer Region. Sie umfasst für einen Naturraum etwa 150 bis 250 Pflanzengesellschaften, von denen aktuell jedoch durchschnittlich nur die Hälfte verwirklicht ist.
Dieses Potenzial einer verborgenen Vielfalt, das je nach Region in einem mehr oder weniger tiefen Dornröschenschlaf schläft, beschreibt gleichzeitig einen Fächer an Möglichkeiten, die vom Bewirtschafter aufgegriffen werden können. Jede Region hat also ihr Potenzial, ihren Fächer an Möglichkeiten, weswegen man die Summe der Möglichkeiten auch als landschaftstypisches Leitbild verstehen kann. Ich verwende den Leitbild-Begriff hier jedoch nicht gerne, da er so aufgefasst werden kann, dass dem Landwirt etwas vorgegeben wird und er selbst nur noch der Ausführende ist. Das ist hier aber nicht gemeint.
Vielmehr präsentiere ich die Palette an möglichen Pflanzengesellschaften als eine Art Vorschlag, mit dem sich die Menschen vom Hof vertraut machen können und die ihnen Anregungen geben, ihre eigenen Wünsche und Träume zu entwickeln, zu ergänzen, zu verändern, zu bereichern. Es hat sich gezeigt, dass diejenigen Pflanzengesellschaften der Kulturlandschaft, die die größte botanische Vielfalt auf sich vereinigen, zu 10 Biotop-Typen zusammengefasst werden können. Diese sind alle in irgendeiner Weise von Pflege und Bewirtschaftung abhängig, sind also keine „natürlichen Ecken “ oder „Wildnis“. Mehr zum Thema „Die 10 Biotope für die lebendige Vielfalt der Kulturlandschaft“.
Eigene Publikationen zum Thema
- H.-Ch. Vahle (2005): Lichtrasen als Brennpunkte zwischen Erde und Kosmos in der Landwirtschaft gestalten. – In: Landwirtschaft und Kosmos. Dokumentarband zur Landwirtschaftlichen Tagung 2005 am Goetheanum. Hrsg.: Sektion f. Landwirtschaft d. Freien Hochschule f. Geisteswiss. am Goetheanum von S.O. Mahlich: 161-167.
- H.-Ch. Vahle (2006): Lichtvegetation in die Landwirtschaft! – In: Van Elsen, Th. (Hrsg.): Einzelbetriebliche Naturschutzberatung – ein Erfolgsrezept für mehr Naturschutz in der Landwirtschaft. Beiträge zur Tagung vom 6.-8. Oktober 2005 in Witzenhausen: 99-108.
- H.-Ch. Vahle (2006): Steigerung der Vegetationsvielfalt im Ökolandbau. – Jahresberichte Forschungsring Biologisch-Dynamische Wirtschaftsweise 2006: 50-52. Darmstadt.
- H.-Ch. Vahle (2007): Landschaft gestalten – Vielfalt der Vegetation steigern. – Lebendige Erde 58 (4): 12-15. Darmstadt.